李英:除了帮熊猫吃竹子,肠道菌群甚至促成新物种!
2019-11-08 15:13 来源:四川省微生物学会
四川农业大学李英应邀参加近日成都举行的“第四届国际农业生物刺激剂大会”演讲录制。
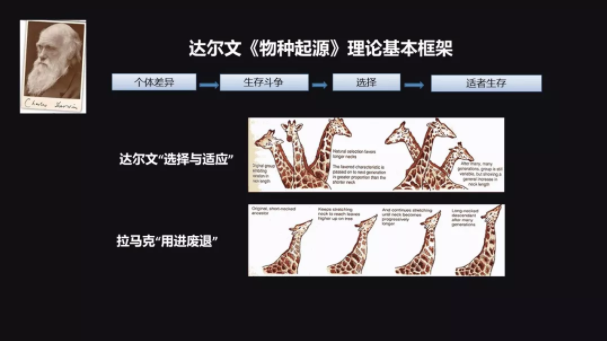
说到进化,我们不得不提到达尔文,他在1859年发表了一个旷世巨著叫做《物种起源》,该书基本的框架就是:第一,它认为自然界的资源是有限的;第二,它认为在群体之间个体是有差异的。
这些差异的结果就会导致个体在竞争有限资源的时候,有些个体更能适应这个环境,那么它就更能生存下来,并且获得更多的后代。
我以一个大家比较熟悉的例子来给大家讲解长颈鹿的问题。为什么长颈鹿的脖子是长的呢?
在达尔文以前,拉马克的“用进废退”学说是比较占主导的。他认为只要是需要的就可以得到,就是说你需要这样的长脖子,你就可以进化出这样的长脖子,而且你上一代经过训练得到的一些技能可以遗传到下一代,也就是所谓的获得性遗传。
而达尔文不这样认为,他认为本来在这个群体里面就有一些长颈鹿的脖子是比较长的,那么它就可以得到更多的树叶,这样能够生存得比较好,能够有更多的机会留下后代。经过这样一代一代地选择,长颈鹿的脖子就变得越来越长。
而按照达尔文的理论,我们所有现存的生物以及已经灭绝的生物都有一个共同的祖先,只是说有一些事件发生得还比较近。
比如说我们看人和黑猩猩在大约600万年前发生分歧而分开了,那我们还能看到我们和黑猩猩有很多很类似的特征,比如骨骼、表情。
但是有一些物种已经分歧了很久了,比如说高等植物、高等动物和细菌,看起来好像没什么关系了,实际上它们仍然是有联系的。
比如,按照“内共生假说”的内容,高等动物的线粒体是来自于一种可以进行三羧酸循环的这样一个细菌;高等植物中的叶绿体是来自于一种可以进行光合作用的蓝藻(蓝细菌)。
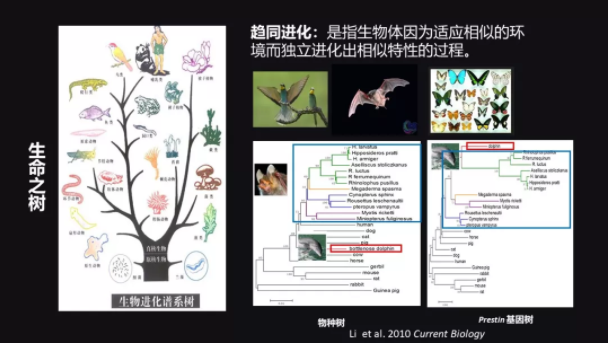
我们可以用一棵“树”比较形象地把所有现存的生物或者已经灭绝的生物都放在这棵“树”上,那这棵“树”上每一个枝之间的距离就代表了它们进化关系的远近。
还有一种比较特殊的情况,在有的时候,有一些生物为了适应相似的环境而独立进化出来相似特征的这样一个过程,叫做趋同进化。
在形态上一个非常典型的趋同进化的例子就是,我们知道鸟、蝙蝠、还有蝴蝶都能飞翔,但是在进化上鸟是属于鸟纲,蝙蝠是属于哺乳动物,它是哺乳动物中唯一会飞的,蝴蝶是属于昆虫纲。这三个进化上很远的东西都进化出来了翅膀,它从形态和功能上都是很类似的,这就是在形态学上的趋同进化。
那在基因进化上有没有这样的现象?也是有的。
我先介绍两个概念,一个叫“物种树”,一个叫“基因树”。能够真实代表这些物种的进化关系的,我们叫它“物种树”。有一些基因,我们用一些分子标记来构建出系统发育树,这个树呢,它未必能够体现物种树,那这个时候我们叫它“基因树”。
我这里也给大家举一个例子,我博士后期间工作的一个小例子。我们发现蝙蝠和海豚其实在进化上距离是比较远的,一个属于翼手目,一个属于偶蹄目。左边这一张图是它们的物种树的关系。
我们发现在 Prestin 基因(这是一个跟听力相关的基因)的基因树上,蝙蝠就跟海豚聚在一起了,那是因为它们都有一个共同的特征,叫做回声定位,它们可以用回声定位来寻找食物,逃避敌害。
这个工作也是在分子进化上一个趋同进化的非常典型的例子。
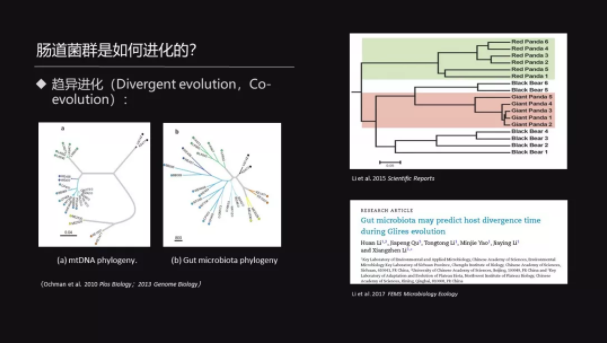
那么我们想问一个问题——菌群是怎么样进化的?跟宿主的基因进化很类似,我们也可以把它简单地归为两类。
一个是趋异的进化——也叫 Co-evolution,也就是说,当菌群的进化的聚类关系体现的是宿主的系统发育关系的时候,我们叫做 Co-evolution。
那么一个比较典型的例子就是,Ochman 他们在2010年的时候发现,菌群的聚类关系和用线粒体 DNA 构成的系统发育树的关系是一致的。
我们的研究也发现,尽管大熊猫和小熊猫它们在食性上是趋同的,但是它们的菌群依然体现了系统发育的关系。
另外,有最近的一项工作甚至发现,菌群的进化和宿主的进化是同步化的,甚至你可以用菌群的进化来预测物种的分歧时间,就是宿主的分歧时间。

另外一种叫做趋同进化。
趋同进化最典型的例子是Jeffrey Gordon他们在2011年发表在 Science(《科学》杂志)上的一篇文章,他们发现哺乳动物的菌群的进化是跟它的食性是一致的。
2014年 Rob Knight 课题组也发现,食蚁的这些动物的菌群进化也是有趋同的现象。
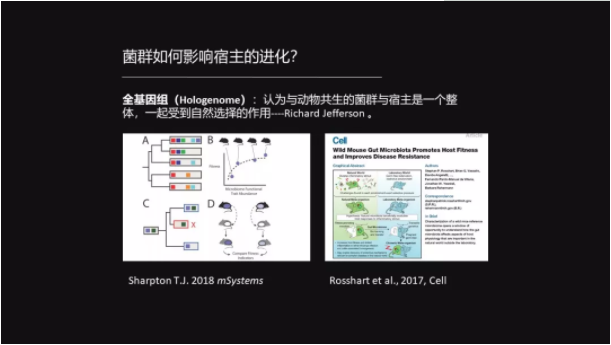
而我们更想知道的是:菌群的进化如何影响宿主的进化?
在1989年的时候 Richard Jefferson 提出来一个观点——全基因组的概念,他认为与动物共生的菌群应该跟宿主是一个整体,它们一起受到自然选择的作用。
当一种菌所具有的功能能够让它的宿主适应新的环境,或者更好地适应原来的环境,那么这种菌的丰度就有可能跟这个宿主的适应度是正相关的。
那这样的例子也挺多的。比如说 Rosshart 他们在2017年发现,如果把野生老鼠的菌群移植到实验室老鼠中,就会提高实验室老鼠的生存率以及它的抗病性。
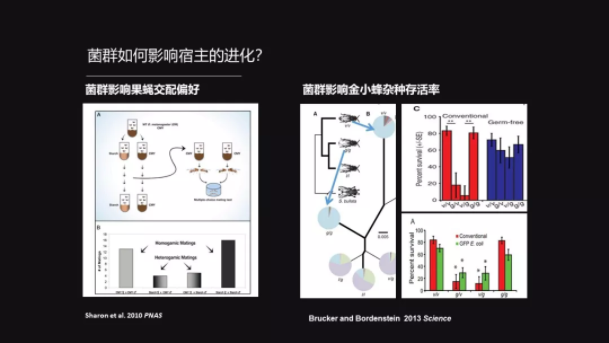
菌群除了能够影响宿主的环境适应性之外,它甚至可以影响物种形成。这里有一个比较经典的例子。
在1989年,Dodd 他们的研究发现,喂一群黑腹果蝇要用两种不同的食物,一种食物是淀粉培养基,另外一种食物是蜜糖培养基。他们发现这些吃不同食物的果蝇在选择配偶的时候,它们倾向于选择那些跟它吃同样食物的果蝇来进行交配。
后来的研究发现,如果把这些果蝇用了抗生素处理之后,让它没有菌了,或者是菌减到很少,那么这种交配的选择就会消失。可以说这种交配的偏好性就是由于菌群导致的,而且交配的偏好性继续下去,就会形成生殖隔离——就是同一个物种的两群就不再交配了。生殖隔离就是物种形成的一个标志。
更直接的一个证据来自于金小蜂的研究,这不是一个人名,是一个寄生蜂。这两种金小蜂其实已经形成了物种隔离,也就是说,它们要么就是交配了以后不能产生后代,要么产生后代而后代不能生存。
但是,有研究表明,如果把金小蜂做一个无菌化处理,那么这种生殖隔离就会消失;同时给无菌化处理的金小峰移植了大肠杆菌之后,那生殖隔离就又会出现。所以说这也是直接地证明菌群影响了物种的形成。

那么菌群怎么样影响宿主对食物的适应?我想大熊猫应该是最合适的一个模型来研究这个问题。
大熊猫是我们国家的国宝,也是世界野生动物保护的一个旗舰物种。它在分类地位上是属于食肉目中的熊超科,目前保护地位是易危级的。
在古代的大熊猫和现代大熊猫相比,它的生存空间要大三倍左右。而且古代大熊猫是杂食性的,一直到了距今5000年到7000年之间,古代大熊猫才跟现代的大熊猫一样变成专吃竹子。
现代大熊猫大概食性里面99%是吃竹子的。我们众所周知,竹子是以低蛋白质、低脂肪和高纤维为特征的,它是一种低营养的食物,而且在大熊猫的基因组里面并没有找到编码可以消化这种纤维素的基因。

那么问题来了,为什么大熊猫要选择吃这种又没有营养,自己又不能消化的食物呢?
有研究表明,有一个基因叫做鲜味受体基因,它是能够让你尝到肉的鲜味的,而大熊猫的这个基因变成了一个假基因所以有人就认为大熊猫为什么要吃竹子而不吃肉了,是因为这个基因变成了一个假基因。但实际上当我们看牛和马这些草食动物,它们的 T1r1(鲜味受体)基因依然是完整的。
因此,这个说法其实是不能解释大熊猫的食性转换的。
另外还有一个研究认为,有一个基因叫 COMT,它是跟食欲相关的。这个研究就认为,大熊猫这个基因上缺失了12个核苷酸,而缺失这12个核苷酸就导致它的这个基因不完整,就影响了它的食欲,从而影响它对食物的选择。
后来我们的研究组也是又找了很多个体来进行了测序,发现并没有缺失之前的报道描述的那12个核苷酸,可能是一个测序的错误或者拼接的问题。
Ok,到目前为止“大熊猫为什么吃竹子”这个问题还是个谜。但无论如何,它已经吃了几千年的竹子。我们就想知道,它是怎么样来适应这种高纤维、低营养的食物的呢?
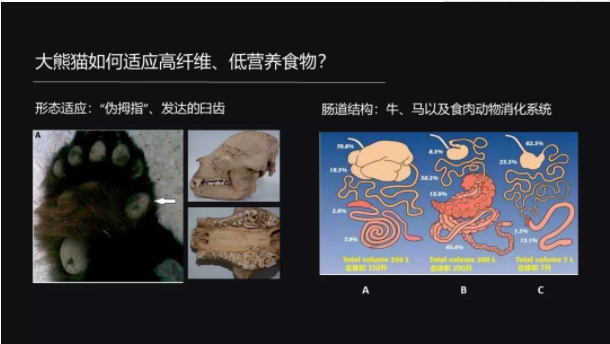
首先从形态上来说,大熊猫的臼齿非常发达,这样发达的臼齿可以帮助它咬碎食物;另外它的手腕上有一个籽骨,这个籽骨就发育成了一个“伪拇指”,这个伪拇指跟其它的五个指头配合,就能让它更好地握紧竹子,这是它形态上的一个适应。
那么它抓到竹子了,到底能不能消化这个竹子呢?我们来看一下它的肠道结构。
跟牛和马相比的话,牛我们知道是反刍动物,有4个胃,可以把食物经过长时间的发酵;而马有很发达的盲肠,也是可以发酵食物。
但是大熊猫依然保留了食肉动物的这种比较简单的肠道,它的肠道很短而且很直,盲肠是退化的,那这样的结构和草食动物是不一样的。

我们很自然地想到:大熊猫的肠道菌群能不能帮助它来消化这些纤维素呢?
2011年,Zhu 等他们通过对15只大熊猫的研究发现,一些梭菌属的菌群是可以降解纤维素和半纤维素的。同时 Xue 等他们在2015年对圈养大熊猫也发现了同样的结果。
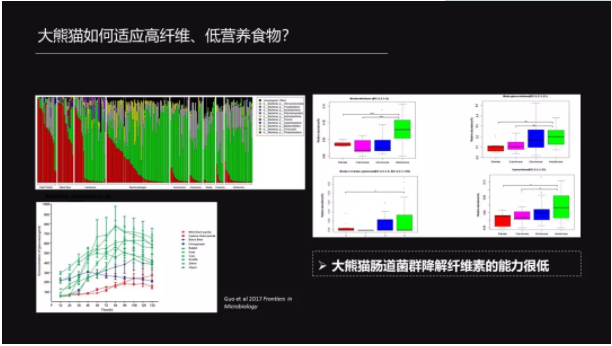
然而我们的研究发现,大熊猫不管是从菌群的组成,还是从它的功能、它的酶活性来看,都是比较类似于食肉动物的——就是它的酶活性是比较低的。
那么在这种情况下,我们认为竹子可能不能提供给大熊猫很好的、足够的能量。
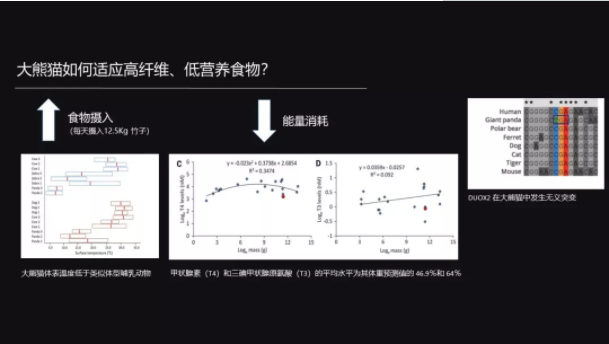
那大熊猫怎么来解决这个问题呢?我们想应该是两个,一个是开源,一个节流。
什么叫开源呢?开源就是增加食物的摄入,一只大熊猫平均每天要花10个小时来吃竹子,每只大熊猫每天要吃掉12.5公斤的竹子。
那节流呢?Nie 等他们发现大熊猫的体表的温度要比同体积的其它哺乳动物的体表温度要低,而且它的甲状腺素 T4 和 T3 的分泌都比其它的哺乳动物要低。
因此,我们综合来看,就是大熊猫是如何来适应吃竹子这样的一个食物的转换?其实它是从形态、肠道微生物以及内分泌综合的一个过程。

环境因素其实是自然选择的一个非常重要的选择压力。
青藏高原是以低温、低氧、高紫外线作为一个极端环境的典型。我们平原人只要快速地上到3000米的这个海拔的时候,大概50%-75%的人会出现或轻或重的高原反应,高原反应轻的人就是被紫外灼伤,然后呼吸困难;那重的可能就是一些很严重的心肺损伤,甚至会危及生命。
但是我们看那些土著的高原动物——牦牛、藏猪、藏羚羊,还有藏族人,它/他们能很愉快地在高原上生活。
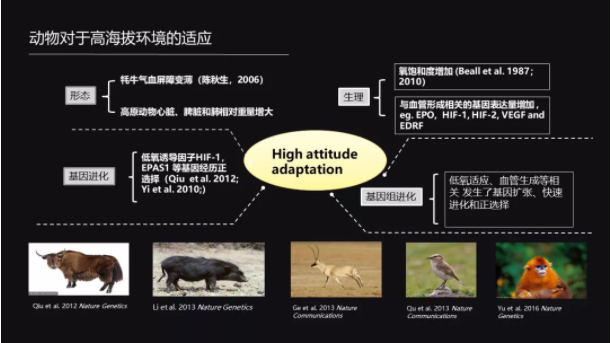
那么它们是怎么样适应这个高原的环境呢?其实科学家从形态、生理和基因等等方面已经找到了很多的证据。
比如说牦牛的气血屏障变薄;比如说高原动物的心脏、肺脏以及脾脏的相对大小变大;比如说跟血管形成相关基因的表达增加。这样的一些变化都能有助于它们来适应这种低氧的环境,而这些变化其实已经深深地烙在了它们的进化里。
从基因组上我们可以看到,它们这些跟低氧适应相关的一些基因经历了正选择或者是经历了基因的扩张。

那么菌群在这个过程中能不能起一些作用呢?答案是肯定的。
Zhang 等他们在2016年对牦牛和藏羚羊的瘤胃的一项工作中就发现,牦牛和藏羚羊这两种高原动物的甲烷排放量比较低;宏基因组的结果也显示,它们的一些基因的 pathway(通路)的丰度也是比较高的,短链脂肪酸的基因丰度也是比较高的。
这样的一些变化是有助于适应这样一个低温低氧的环境的。

我们小组的研究也发现,藏猪和藏族人有比较相似的菌群,而且我们通过代谢组也能发现他们的一些代谢通路,比如说,脂肪酸代谢、氨基酸合成代谢这样的一些通路是富集的,并且像脂肪酸这些代谢产物也是比较多的。
那这样的一些东西可以让他们迅速地获得能量,就是能够适应高原环境的这种低温的特征。
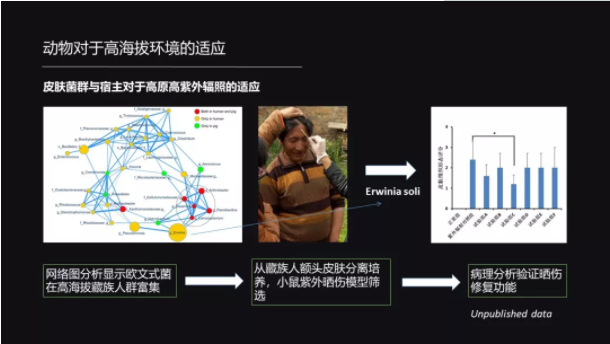
此外,皮肤上的菌群对于宿主适应高原的高紫外线也是有帮助的。
我们用高通量测序的方法发现,在藏族人里面欧文氏菌是比较富集的。于是我们就从藏族人的额头开始分离这种欧文氏菌,分离了几百株这样的菌,然后我们又建了小鼠晒伤的一个模型,然后用这个菌来涂抹。
结果我们就发现,有那么一株菌能够显著地修复这种小鼠的晒伤。
所以从以上这些例子就可以看到,肠道菌群或者皮肤菌群对宿主、对高海拔环境的适应也是有帮助、有贡献的。

那我说了那么多,可能有人会问我:“研究菌群的进化到底能给我们什么样的启示呢?”

首先我们看,对于野生动物,我们说,现在地球上已经经历了5次大范围的生物大灭绝,每一次的生物灭绝都有一些类群消失殆尽。
而我们现在正在处于第6次生物灭绝的一个开端,这一次我们将会失去很多的哺乳动物和鸟类,失去的生物多样性、失去的物种会让我们失去非常宝贵的自然资源,而更重要的是,它会降低我们整个生态系统的稳定性。所以我们要保护遗传多样性,保护生物多样性。
我们以前的工作都是集中在基因方面,比如说我们保护遗传多样性。最近的研究有不少工作集中在菌群上,包括我们自己的研究。
我们也发现在迁地保护的过程中——迁地保护就是说对一些野生珍稀动物,我们给它建一些繁育中心,建一些动物园,然后把它迁到繁育中心来进行保护,来让它在这生活。在这样的一个过程中野生动物的菌群损失了很多。
以我们的大熊猫的研究为例,圈养大熊猫的菌群只有野生大熊猫 OTU 的26%,损失掉的那些菌群可能跟大熊猫的环境适应性、跟大熊猫的消化以及疾病的抵抗有关系。
所以我们得到了这么多的信息和知识,也不得不思考一些问题:菌群作为动物的第二基因组,我们是不是应该保护它?我们如何来保护它?我们是不是在训练、 野化、放归这些动物的过程中也应该训练它们菌群的适应性?
我想这些问题是一些新的问题,也是我们值得思考的一些问题。

其次我们现在的家养动物,比如说家猪、家牛和家兔等等,大概都是在距今1万年,是由我们人为驯化而来的,是从野生动物驯化而来的。
那么在驯化的这个过程中,如果我们是把它当宠物,我们就根据自己的喜好或者说我们的需求来给它们压力。比如说奶牛,我们想让它产生更多的奶,这样一个压力就给它,持续不断地给这样一个压力,并且这个压力是一个很大的选择压力,在这样的一个选择压力下,就形成了现在我们这样的家养动物。
现有的一些野生动物,比如说像野猪还存在,它的种群还比较繁盛。研究也表明,这些野生动物具有疾病耐受性比较强、比较耐粗饲这样的一些特征,那我们可以利用这样的一些特征跟它进行一些杂交。
野猪跟现代的家猪进行杂交,杂交完了以后叫特种野猪,那特种野猪会兼具这两种猪的一些优良品质。
我们的研究发现,野猪菌群的多样性是高于其杂交后代的,那就是说我们在家养动物驯化的这个过程中同样丢失了很多微生物。
我们的课题组也在致力于把这些微生物分离并培养出来,目的是想找到一些能够耐粗饲的、能够提高免疫力的这样的一些菌,作为潜在的益生菌。
为什么要做这个事情?因为大家知道,我们现在养殖业里面用的抗生素是比较多的,而这个已经成为一个很重大的问题。
那为什么抗生素的使用会成为一个很重大的问题?我们从菌群本身的进化上其实是不难理解的。
第一点,细菌的世代时间是比较短的。以大肠杆菌为例,大概是30分钟一个世代,而且它的基因组是很紧凑的,没有内含子。所以你只要有一个碱基突变,就有可能改变这个氨基酸,而改变氨基酸,就有可能改变功能。
第二点,很多抗生素的抗性基因是在质粒里面的,质粒是很容易横向传播的。
第三点,我们宿主给它很多的选择压力,比如我们宿主的基因型,宿主给它提供的营养,以及宿主的用药等等都会很快地影响这个菌群的进化。
因此菌群的进化是要比我们快很多。
在2006年的时候,欧盟已经提出来限制抗生素的使用,我们国家在2018年也颁布了限抗令。在这样的一个国家需求之下,我觉得解铃还需系铃人——就是微生态的应用可以是代替抗生素的一个非常有前景的替代物。

最后,我想用一句话来结束我的演讲。著名的进化生物学家 Dobzhansky 说过这样一句话:“Nothing in Biology Makes Sense!Except in the Light of Evolution!”
我也希望我们能在进化之光的指引之下,能够一层一层揭开菌群的神秘面纱,最终利用它来让我们人类和动物生活得更健康。
上一篇:第十六届复旦大学世界校友联谊会--生物医药峰会
下一篇:第四届国际农业生物刺激剂大会落幕 川大生命科学院孙群等作分享
TOP排行
- 你好,微朋友——四川省微生物学会携手四川师范大学成功举办科普开放日活动
- 联学共建促科普 深地科学筑梦想——四川省微生物学会专家科普讲座走进西昌怀远学校
- 四川省微生物学会参加首届中国酒业产教融合高质量发展大会
- 中国微生物学会第十四届地质微生物学学术研讨会拟于4 月在兰州召开
- 成都生物所胡军华团队合作揭示数据提质助力中国生物多样性保护再续佳绩
- 微生物所团队合作揭示镰孢菌在高原农田生态系统的分布格局并定名描述“墨脱镰孢”等藏南新种
- 进展 | 赵瑞琳团队系统解析裸盖菇素生物合成基因簇的起源与演化机制
- 微生物所叶健团队发表植物病毒虫媒传播评述文章
- 微生物所叶健团队揭示机械力受体PIEZO1介导的植物力学抗虫免疫新机制
- 四川省微生物学会跨界搭台 《梅花新韵》,奏响科艺融合新乐章

